
文|面包夹知识
编辑|面包夹知识
«——【·前言·】——»
全球变暖已成为一个迫在眉睫的世界性问题,减排和增汇是实现大气中CO2减少的必然途径。

目前世界各国对于减排的手段层出不穷,而对于增汇的关注略显不足。海洋是地球上最大的活跃碳库,作为最大的碳汇途径,每年可吸收约30%的人为释放的CO2。
2018年人类活动排放的CO2总量约为(11.5±0.9)Gt,其中约(2.6±0.6)Gt被海洋吸收。

海洋巨大的碳吸收能力对于减少大气中的CO2和减缓全球变暖具有重要作用,海洋碳汇是指红树林、盐沼、海草床、浮游植物、大型藻类、贝类等从空气或海水中吸收并储存大气中CO2的过程、活动和机制。
渔业碳汇是指促进水生生物通过渔业生产活动吸收水中CO2,通过收获水生生物来去除水中的碳或通过生物沉积作用将其沉降于水底的过程和机制,故也被称之为“可移出的碳汇”。

这一理念充分显示了水产养殖的食物供给和生态服务两大功能,多营养层次综合养殖模式是一种低碳、生态、高效的水产养殖模式。
生物碳汇既可以改善生态环境增加渔民收入又可以人为增加,能够较好的体现出碳汇渔业的新理念,从而有助于中国低碳经济的可持续发展。
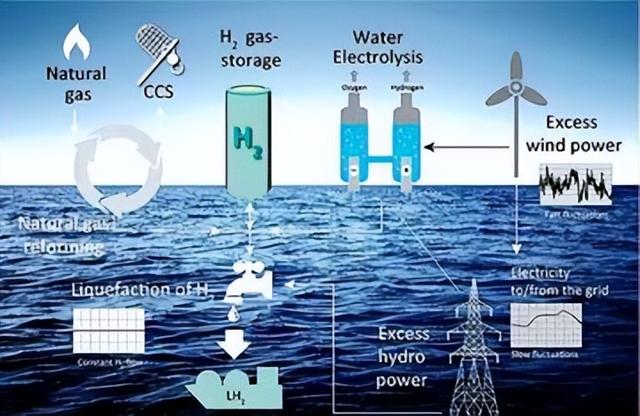
我国是世界上最大的海水贝类和藻类养殖生产国家,据《中国渔业统计年鉴》统计数据显示,2021年我国海水贝、藻养殖产量为1798万t,约占我国海水养殖产量的81.3%,约占世界海水养殖产量的50%。
研究表明,2018—2020年间我国贝、藻养殖总碳汇量高达659万t,相当于每年造林87万hm2,体现出巨大的碳汇功能。

贝藻IMTA模式既能更好地体现水产养殖的生态效益,又能发挥贝、藻的碳汇功能,是碳汇渔业的代表性发展模式。
大力发展海洋碳汇渔业IMTA模式,突破改进海水养殖业的关键技术,解决目前养殖模式中存在的问题,进行贝、藻等多种生物的人工增养殖,积极拓展改革生态系统养殖模式,对于低碳经济发展和实现“碳中和”目标具有重要意义。

«——【·大型藻类碳汇过程及机制·】——»
大型藻类碳汇是指利用大型藻类从空气或海水中吸收并固定CO2的过程、活动、机制和能力,由大型藻类沉积物碳汇和大型藻类植物碳汇组成。
大型藻类在生长过程中,通过光合作用吸收海水中的溶解性无机碳,和CO2生成颗粒有机碳和溶解有机碳。
POC在生物泵的作用下或沉积并掩埋在海底,或通过直接沉积输送到深海,或通过天然食物链成为其他生物的食物来源。

除此之外,大型藻类还通过自身产生碎屑方式沉积至海底形成长期难降解的稳定性碳以及通过收获大型藻类本身来形成海洋碳汇。
大型藻类产生的DOC通过微生物碳泵在微食物环作用下,进入食物网或形成惰性溶解有机碳而长期驻留在海水中。
藻类在生长过程中还通过吸收溶解在海水中的无机营养盐,使得海水的碱度升高、CO2分压降低,在溶解度泵的作用下促进海水对空气中CO2的吸收,实现海洋碳汇功能。
大型藻类具有很高的固碳能力,研究表明大型海藻能够在短时间内积累大量的碳储存,并能够在碳循环中作为生物碳汇发挥关键作用。
估算在全球范围内大型藻类可以每年封存碳约173Tg,其中约有90%是通过碳沉降迁移至深海实现长期固碳。
例如,在沿海某研究员,通过推断以及数据调查表明,养殖的大型海藻每年约吸收124.05万t碳。

而估算在1999—2008年间,平均每年通过收获养殖海藻从我国近海被利用的碳量为379万t,其中有120万t的碳通过捕捞移除。
而在我国的南海,一些研究专家研究表明,大型藻类的碳封存途径有73%是通过POC途径,若对大型藻类,进行干预性保护可能将能够去除全球10%左右的碳。

估算出大型藻类约有43%的初级生产力以生物体、DOC和POC的形式进行了碳排出,其中约有24%被封存在深海沉积物中。
大型藻类的固碳能力也会因种类的不同而有所差异,比较了3种海藻收获后各自所属养殖场表层水的pH变化。

其中海带显示出较强的CO2吸收能力,其养殖区域内的pH上升了0.10,龙须菜和紫菜的pH分别上升了0.04和0.03,说明大型海藻在缓解海洋酸化方面也有重要作用。
在1999—2012年间,全国海藻年均固碳量为418.5万t,其中海带的年均固碳量(264.5万t)最高,裙带菜(32.3万t)和紫菜(22.4万t)分列第二、第三,且海藻固碳量跟产量呈正相关关系。

«——【·滤食性双壳贝类碳汇过程及机制·】——»
双壳贝类碳汇是指利用双壳贝类从空气或海水中吸收并固定CO2的过程、活动、机制和能力,由贝类沉积物碳汇、贝类贝壳碳汇和贝类软组织碳汇组成。
滤食性双壳贝类通过过滤、呼吸、钙化和生物沉积等过程与浮游植物、颗粒有机碎屑、海水碳酸盐系统、沉降和掩埋过程相互作用,对碳的生物地球化学循环产生重大影响。

滤食性双壳贝类在成长过程中可以通过大量滤食浮游植物及有机物碎屑等POC供自身的一系列生理活动而增加生物体含碳量,并且能够以粪便颗粒的形式将未使用的有机碳沉降于海底。
据研究,葡萄牙南部沿海的养殖贻贝(Mytilusedulis)每形成1gCaCO3可释放0.29gCO2。

例如,美国科学家发现,波罗的海沿岸的养殖贻贝,每形成1gCaCO3则释放高达0.385g的CO2,这表明CO2的释放量可能跟海域环境有一定关系。
我国的研究人员也表明,在山东半岛桑沟湾养殖的双壳贝类通过钙化作用固定的无机碳要高于通过呼吸作用释放到海水中的CO2,表明双壳类动物是碳汇而非碳源。

在意大利北亚得里亚海开展的蛤蜊(Ruditapesphilippinarum)和贻贝混养试验表明,蛤蜊和贻贝通过壳的形成分别可以永久捕获254g和146g的CO2,此外分别有22g和55g的CO2用于呼吸排放,蛤蜊和贻贝养殖可被视为碳汇。
研究表每年通过从大亚湾收获葡萄牙牡蛎(Crassostreaangulata)贝壳累积移出的碳量为258g/m2,而因呼吸和钙化释放的碳量为502g/m2,该养殖活动整体表现为碳源。

科研人员,通过代谢过程和贝壳形成释放的CO2量,比海洋环境中贝壳中封存的碳量多,贝壳是CO2的净来源。
关于养殖贝类是碳源还是碳汇仍然存在争议,近年来的诸多研究则倾向于认为贝类碳汇不是作为一个单独的过程,而是被整合到整个海洋生态系统中,例如其与浮游植物种群。

悬浮POC和DIC相互作用的耦合可以显着改变CO2循环。贝壳具有较高的固碳速率并且固碳占比较大。
从1999—2014年的研究数据发现,通过贝类养殖每年有(0.90±0.10)Mt的碳被从海域中移除,其中贝壳固碳量(0.69±0.06)Mt。
研究表明在牡蛎养殖期间贝壳固碳量约为每年1.31Mt,约占总去除碳量的83.96%。

据测算,山东半岛桑沟湾养殖扇贝(Scallop)的固碳速率为每年3.36t/hm2,明显高于自然水域“蓝碳生物”的固碳速率。
值得重视的是,有研究表明,当水体中pH为7.3时贝类的钙化率接近于0,贝类将不能合成贝壳,这对贝类的生存和其碳汇功能都非常不利,所以有必要采取措施防止海洋酸化。

目前已有很多关于贝类碳收支的研究,基于能量收支分配模型“使用碳=移出碳+储存碳+释放碳”,栉孔扇贝(Chlamysfarreri)摄食的有机碳中有17.24%供给呼吸排碳,有66.09%供给粪便和代谢排碳,有16.67%供给自身生长。
例如,美国科学家认为,在对长牡蛎(Crassostreagigas)一个生长周期(360d)的研究发现,单个长牡蛎可吸收水体中14968mg碳。

其中有3600mg通过贝壳固碳,有430mg通过软体部固碳,有3322mg通过生物沉积物固碳。
对菲律宾蛤仔的碳分配发现,约有46.2%的碳沉降至海底,约有40.6%的碳用于呼吸和钙化并重新释放进入水体,约有13.2%的碳通过收获移出。

而在菲律宾威尼斯泻湖对蛤蜊的一个生长周期进行了研究,发现有124kgCO2用于贝壳固碳,有88kgCO2用于软体固碳,还有额外的56kgCO2用于呼吸排出。
不同种类的贝类其固碳能力也有所不同。
有研究分别对缢蛏、河蚬和光滑河兰蛤的有机碳分配做了较系统的研究,指出缢蛏固碳占比49.95%,河蚬固碳占比62.60%,光滑河兰蛤固碳占比为76.92%。

例如,在意大利萨卡迪戈罗泻湖对蛤蜊的生命周期评估中发现,每1t蛤蜊在其生长周期结束时可净封存444.55kgCO2,相当于固碳占比44.46%。
在2003—2019年对我国贝类年均固碳量计算中,贡献率最高的主要贝类物种为长牡蛎、菲律宾蛤仔和栉孔扇贝,对年固碳能力的平均贡献率分别为39.36%、22.32%和14.44%。

«——【·贝藻养殖增汇模式·】——»
多营养层次综合养殖(IMTA)模式是在一个系统中养殖来自不同营养层的水产养殖物种的养殖技术,是一种实现环境和经济可持续性的做法。
在贝藻IMTA系统中有机提取单元(主要为贝类,可过滤颗粒有机物)和无机提取单元(主要为大型海藻,可吸收无机营养盐)可以吸收来自饲饵式鱼虾类的有机和无机颗粒废物,降低养殖对环境的危害。

研究显示,IMTA技术可增加水产养殖动物和海藻的产量,同时减少其他环境污染物的产生。
IMTA的起源可以追溯到数千年前,现代IMTA的概念最初是由19世纪70年代提出的,他们将不同营养级别的生物结合在一起,实现了IMTA的基本概念。

此后,世界范围内陆续设立了许多IMTA养殖基地,其中我国的桑沟湾基地和獐子岛基地是世界上最大的IMTA养殖基地。
目前我国海水养殖的IMTA模式有贝-藻、贝-鲍等混养模式。

欧美国家(如意大利、加拿大以及智利等)海水养殖的IMTA模式主要包括鱼-贝、鱼-藻、鱼-参等混养模式。
大量实践证明IMTA养殖模式是在经济效益与生态效益上比单一品种更优的养殖模式。
比起贝、藻单养,贝藻混养能够更有效地吸收并利用CO2。

藻类既能够吸收贝类新陈代谢所释放的CO2及有机营养盐等物质,还可以降低海水pH,阻止海洋酸化并有利于贝壳的生长。
贝类在生长过程中可以通过滤食浮游生物及有机碎屑等净化水体,使藻类能够更好的进行光合作用,还可以避免浮游植物与藻类竞争营养盐,为藻类提供良好的生长环境从而促进对CO2的吸收。

通过贝-藻的相互作用,整个综合养殖系统的碳汇功能得到了极大的改善。
海带和皱纹盘鲍(Haliotisdiscushannai)单养每年移除的碳量分别是4397.5kg/hm2和1808.3kg/hm2,而皱纹盘鲍和海带混养移除的碳量可高达12311.9kg/hm2,产生的经济价值每年可达13500元/hm2。

通过贝藻产量增长趋势推测,到2025年我国贝藻养殖吸收的CO2可达606.17万t。
研究表明,养殖大型海藻可以提高牡蛎贝壳的钙化速率,使牡蛎壳生长速度提高40%,从而提高固碳速率。

在瑞典西海岸种植海带和紫贻贝、的试验发现,与单一栽培相比,在贝藻混养模式下海带的生物量和叶片长度显著提高,且海带上的附生植物减少了一半以上。
在IMTA模式下可开展不同混养物种的相关研究,比如扇贝和海带的混养、牡蛎和龙须菜的混养。

也可以对于同一种混养物种而湿质量比例不同进行研究,比如牡蛎和海带分别以湿质量比为1:1、2:1的比例进行培养研究。
对厚壳贻贝(Mytiluscoruscus)和龙须菜在按照湿质量比例为1:0.15以及1:0.30混合下培养发现,海水中的最终CO2浓度分别比比例为1:0.45时分别多5.5倍和2.2倍。

在山东半岛桑沟湾进行贻贝和海带混养的围隔试验,发现贻贝和海带比例为2:1、2:2、4:2和4:3时具有最佳净碳汇功能,且对环境的负面影响最小。
在我国南方大亚湾也做了类似的研究,结果表明牡蛎和龙须菜以4:1的比例混养时,可有效利用海水中的DIC,并进一步增加海洋的碳汇。

室内对海带、牡蛎和龙须菜分别做了生态模拟试验,结果表明大型海藻占比越高碳汇效应越强。
当大型海藻与牡蛎的比例在7:25以上时,该体系表现为CO2的强汇并且能有效降低水体富营养化。

«——【·结语·】——»
我国是全世界最大的水产养殖产业国,海水养殖面积和产量多年稳居世界第一,贝、藻养殖碳汇持续增长,渔业碳汇潜力巨大。
而且近海水产养殖增汇可缓解海洋酸化、阻止全球气候变暖、促进低碳经济可持续发展。

然而气候、生态和养殖空间等各种因素均会影响贝、藻的养殖结构和规模,是预测碳汇的不确定因素。除了可移出的碳汇,“看不见”的碳汇也不容忽视。
水产养殖环境中的碳封存途径主要包括BP介导POC向深海乃至海底传输,MCP过程形成惰性有机碳,以及碳沉积于深海中等。这都应包含在水产养殖碳汇定量评估的范围之内。

目前关于海水养殖中碳汇边界的定义、测汇标准以及“看不见”碳汇的计算方法都还不是很明确。
不过增强渔业企业和渔民积极发展渔业碳汇的动力,共同推进渔业碳汇的发展,才能更好的贝藻养殖。
